Introduction
The combination of dietary excesses and sedentary lifestyle promotes weight gain and obesity, with all the consequences that derive from them. In the ongoing fight against obesity and related disorders, several strategies have proven successful in activating the thermogenesis of brown adipose tissue (BAT) and the energy expenditure of the whole body (EE24h) through the activation of the sympathetic nervous system (see PPG Stress Flow device - Biotekna), such as physical activity, exposure to cold for short periods of time, the use of certain classes of chemicals, including thermogenic agents, supplementation with specific vitamins and minerals and adequate nutrition (see TAM: Alkalizing and Metabolic Therapy for the recovery of vague and nonspecific symptoms - MUS).
Thermogenic agents
Thermogenesis is a process whereby food calories are transformed into thermal energy, which is essential for body homeostasis, i.e. for thermoregulation, the maintenance of metabolism and body weight control. Several thermogenic agents have been studied including caffeine, p-synephrine (bitter orange extract), capsaicin and chlorogenic acid (green coffee bean extract) [29].
Capsinoids
Capsaicin and its non-pungent analogues (capsinoids) are substances that increase energy expenditure and promote the reduction of body fat; in particular, capsinoids activate brown adipose tissue (BAT) thermogenesis and whole-body energy expenditure (EE24h) [30]; to confirm this, in a study carried out on healthy men aged 20-32 years, the measurement of body energy expenditure and skin temperature increased after oral intake of capsinoids (9 mg) due to BAT activation [31,34]; the mechanism of action through which the brown adipose tissue (BAT) is activated is related to the fact that capsinoids activate the TRPV1 channel located in the upper digestive tract, which leads to an increased sympathetic nerve activity towards the BAT.
Caffeine
Caffeine has been shown to increase energy expenditure with a dose-dependent effect and, despite some conflicting data, it is generally accepted that caffeine effectively stimulates the central nervous system (impulsive stimulation of the sympathetic nervous system) and increases metabolic rate [33].
Exposure to cold
Brown adipose tissue (BAT) is activated by acute exposure to cold; as this type of adipose tissue regulates energy expenditure and body fat, it is a promising target in the fight against obesity and related disorders [32]. In a study comparing the use of substrates during physical activity performed in cold and warm environments, respectively, it was shown that total energy expenditure and fat burning during exercise was higher in a cold environment [35].
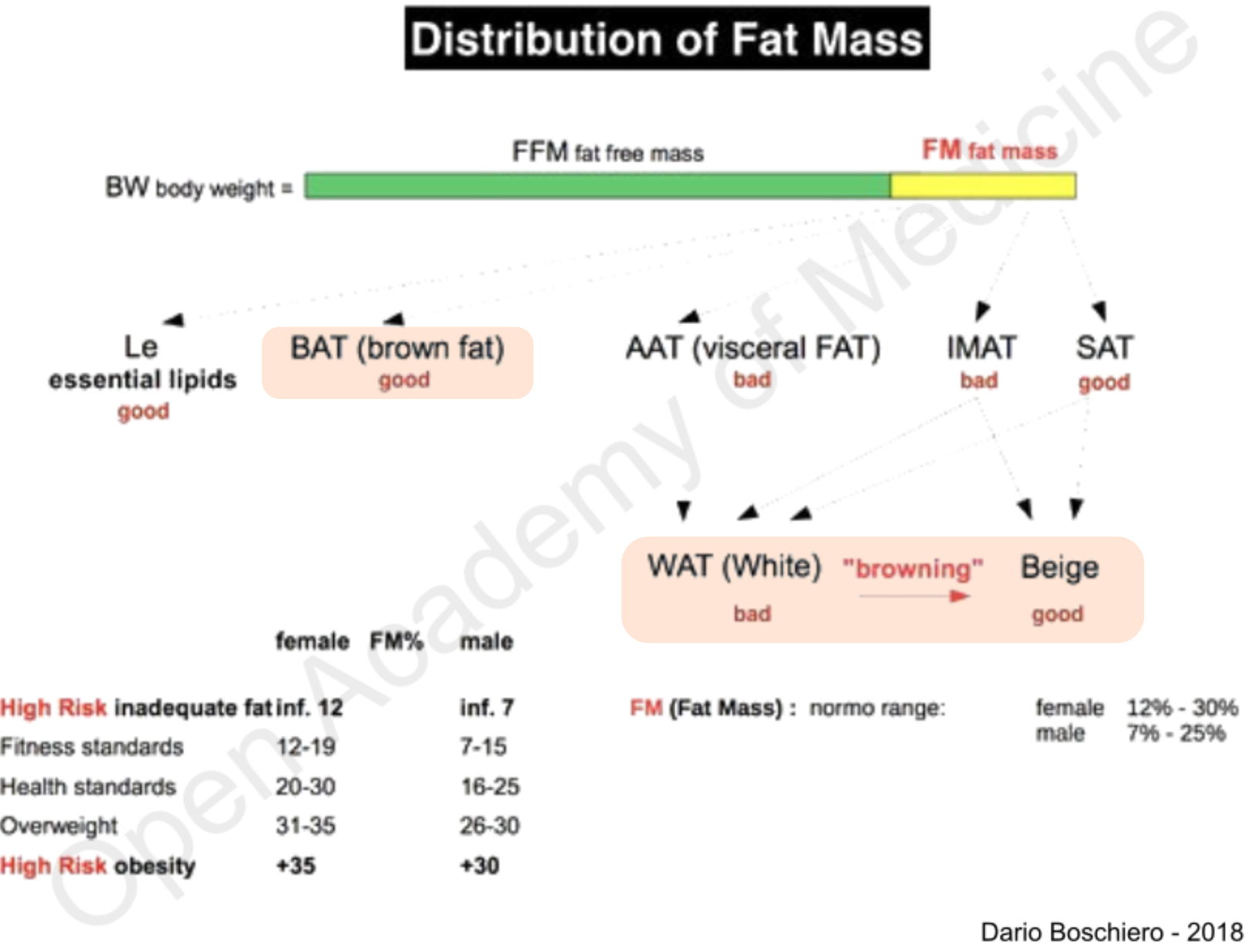
HIIT
Recent research studies clearly show that High Intensity Interval Training (HIIT) may be more effective in reducing subcutaneous fat, abdominal fat and IMAT than other types of exercise. Regular HIIT activity has been shown to significantly reduce insulin resistance and increase the ability of skeletal muscle mass to oxidize fatty acids [36]. In addition, physical activity stimulates the production of irisin, a hormone capable of triggering a molecular mechanism known as “browning effect”, i.e. the transformation of white adipose cells to beige, and, especially in the case of HIIT workout, it leads to an adequate activation of the sympathetic nervous system; it has also been shown that exercise in combination with exposure to cold is essential to stimulate BAT.
Vitamins and energy metabolism of carbohydrates, lipids and proteins
Vitamins play an essential role in reactions that are involved in the metabolism of proteins, carbohydrates and lipids: lipid metabolism is affected by vitamins such as vitamin B1 (thiamine) [1] as well as by folate and vitamins B6 and B12; a deficiency of these vitamins has an atherogenic effect resulting in a significant increase in the lipid accumulation in the aortic adventitia [2]; studies on the activity of vitamin B6 have revealed that high-dose administration of pyridoxine in individuals who followed a diet high in fat and cholesterol leads to a decrease in the concentration of lipids [3,4,5]. Riboflavin (B2) is converted into coenzymes that are involved in the synthesis of fatty acids, while vitamin C is able to significantly decrease serum cholesterol and promote the transformation of cholesterol into bile acids [6,7]. Niacin can be very effective in reducing LDL, cholesterol and triglyceride levels and increasing HDL values: its administration in combination with lipid-lowering drugs (e.g., bile acid sequestrants) has been shown to reduce the incidence of cardiovascular events and slow down the progression of coronary lesions [8,9]. B vitamins are also essential for the metabolism of carbohydrates, as they help transform carbohydrates into energy (ATP) [10]: a deficiency of vitamin B1 (thiamine) causes complications similar to those that occur in metabolic syndrome, such as endothelial dysfunction. The administration of thiamine, in deficient subjects, can prevent the formation of harmful byproducts of glucose metabolism, reduce oxidative stress and improve endothelial function [11], while the administration of vitamin B12 lowers blood sugar. B vitamins are also involved in protein metabolism: many amino acid reactions depend on the presence of vitamin B6, because this vitamin is involved in the transport of amino acids to the cells, it supports the production of amino acids along with other enzymes and is involved in the synthesis and metabolism of choline, methionine, serine, cysteine, tryptophan and niacin [12,3]. Folate is another essential nutrient involved in many metabolic processes including the production of amino acids and the synthesis of nucleotides (DNA) [14].
Minerals and metabolism
Among the minerals that are involved in the metabolism of carbohydrates there are zinc [15,16], copper and chromium. Zinc deficiency adversely affects glucose utilization: its reduced dietary intake during pregnancy affects fetal development and carbohydrate metabolism. A diet low in copper causes a reduced insulin response, thus possibly affecting the use of glucose by peripheral tissues and the release of insulin [17,18]. Chromium is an essential nutrient involved in the metabolism of both carbohydrates and lipids: research studies in this area show that this mineral is able to improve insulin sensitivity and is effective in reducing blood lipids [19,20,21].
Other compounds with energy action
Coenzyme Q10 (CoQ10) is an organic molecule that is useful for energy production [28]; its deficiency causes both respiratory chain disorders, leading to changes in energy metabolism, and the decrease in the mitochondrial membrane potential, which in turn induces apoptosis. L-carnitine is another important factor in cellular energy metabolism. It is spread throughout the body, but its main concentrations are located in the most active metabolic tissue, such as the heart and the skeletal muscle. L-carnitine is essential for the transport of long-chain fatty acids across the inner mitochondrial membrane up to their site of oxidation, where it will produce energy in the form of ATP; one of the most important consequences of carnitine deficiency is the alteration of the metabolic pathways that lead to the production of energy [22,23,24].
Glycemic load control
A lifestyle that includes a low-glycemic load diet has been shown to improve metabolic risk profiles in men and women: there is an increasing number of studies reporting how low glycemic load diets are able to prevent diabetes and cardiovascular diseases [6,27]. The maximum energetic caloric load should occur between 6 a.m. and 9 a.m., when the acrophase of cortisol coincides with the maximum activity of thyroid hormones (see TAM: Alkalizing and Metabolic Therapy for the nonspecific recovery of vague and nonspecific symptoms - MUS); this strategy is useful both to better face the loads throughout the day and because the stimulus to insulin secretion triggers testosterone release and helps stimulate the daytime peak of SNS activity [37].
Conclusions
An appropriate supplementation and a correct lifestyle that includes a proper diet and specific regular physical activity combined with exposure to cold for short periods of time and the use of certain classes of chemicals such as thermogenic agents, are all effective tools to reduce the fat mass. When there is a need for a combination of these strategies to affect the lipid, glucide and protein metabolism, Melcalin DIMET provides an effective support thanks to B vitamins (B1, B2, B6 and B12, along with pantothenic acid and niacin) and folic acid, as well as minerals such as iodine, magnesium and chromium, which target thyroid function, muscular and nervous function and glucose metabolism respectively, while Melcalin Nimet combines the properties of Coenzyme Q10 and L-carnitine with those of minerals such as copper and zinc, involved in cellular respiration and DNA and RNA replication reactions; this formulation is also useful to ensure sufficient energy production, feel less tired and improve the skeletal muscle mass (see advanced analysis of body composition and osteosarcopenic obesity - BIA-ACC Biotekna).
Authors: Dario Boschiero - Date: 26/11/2020
Attention: these contents can be freely used for personal learning purposes only. The use is regulated by Law No. 633/1941 and subsequent amendments, as well as by the copyright and patent legislation in force. Any use for commercial and profit-making purposes is forbidden.
References




